
988368
Wed, Jul 8, 2026

Volume 15, Issue 2 (June 2017)
Iranian Rehabilitation Journal 2017, 15(2): 149-154 |
Back to browse issues page



Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Honarpour A, Mohseni M, Ghavidel Hajiagha S, Irani S, Najmabadi H. Investigation of the Relationship Between a Genetic Polymorphism in ACTN3 and Elite Sport Performance Among Iranian Soccer Players. Iranian Rehabilitation Journal 2017; 15 (2) :149-154
URL: http://irj.uswr.ac.ir/article-1-715-en.html
URL: http://irj.uswr.ac.ir/article-1-715-en.html
Asal Honarpour1 
 , Marzieh Mohseni1 , Siamak Ghavidel Hajiagha2 , Shiva Irani3 , Hossein Najmabadi *1
, Marzieh Mohseni1 , Siamak Ghavidel Hajiagha2 , Shiva Irani3 , Hossein Najmabadi *1
, Marzieh Mohseni1 , Siamak Ghavidel Hajiagha2 , Shiva Irani3 , Hossein Najmabadi *1
1- Genetic Research Center, University of Social Welfare and Rehabilitation Sciences, Tehran, Iran.
2- Department of ??? , School of ??? , Islamic Azad University, Tehran Medical Branch, Tehran, Iran.
3- Department of Biology, Faculty of Basic Sciences, Science and Research Branch, Islamic Azad University, Tehran, Iran.
2- Department of ??? , School of ??? , Islamic Azad University, Tehran Medical Branch, Tehran, Iran.
3- Department of Biology, Faculty of Basic Sciences, Science and Research Branch, Islamic Azad University, Tehran, Iran.
Full-Text [PDF 518 kb]
(3048 Downloads)
| Abstract (HTML) (7912 Views)
Full-Text: (2414 Views)
1. Introduction
The α-actinin-3 structural protein is a fast skeletal muscle protein that is encoded by the α-actinin-3 gene (ACTN3, MIM#102574; chromosomal location: 11q13-q14) [1, 2]. ACTN3 has attracted considerable attention due to a common nonsense polymorphism (R577X) that influences muscular performance [3]. A loss-of-function nonsense mutation (C>T) at nucleotide position 1747 in the ACTN3 coding sequence is responsible for the transition of an arginine (R) to a premature stop codon (X) at residue 577 (R577X; dbSNPrs1815739).
This single nucleotide polymorphism is responsible for the distribution of two different types of ACTN3, both of which are common in general population [4]. The 577R allele is the normal, operational version of this gene, whereas 577X allele contains a changed sequence that prevents the production of a complete functional α-actinin-3 protein [5–7]. It is noteworthy that α-actinin-3 has relatively specialized expression and is limited to the fast, glycolytic muscle fibers that are responsible for quick force generation [6].
It is estimated that 16% of the individuals worldwide, or approximately one billion people, are deficient in α-actinin-3. Our previous study conducted on Iranian normal individuals showed that the allelic distribution of ACTN3 in Iranian population is closer to its distribution in the Caucasian population with 11% homozygosity for the common R577X polymorphism; this is concurrent with the route of migration of the ancient human population from the Iran Plateau toward Europe. ACTN3 knockout mouse model analysis has shown that α-actinin-3 deficiency brings about a “slowing” of fast fiber attributes conjoined with a dramatic fall in grip strength, muscle mass and fast 2B fiber size, a rise in muscle aerobic capacity, and slowing of muscle contractile properties [8].
In an Australian athlete cohort [9], highly significant associations between ACTN3 genotype and athletic performance was revealed. Both male and female elite sprint athletes had significantly higher frequencies of the 577R allele than that of control participants. Furthermore, in a number of studies conducted on European, Israeli, and North American athletes, it has been shown that 577XX genotype is underrepresented in sprint athletes and slightly overrepresented in endurance athletes compared with nonathletic controls [10–15]. These data, in addition to other recent studies, provide strong evidence that α-actinin-3 deficiency is detrimental to sprint and power performance.
The effect of ACTN3 genotype has primarily been studied in elite athletes as its effect on muscle can be most readily discernible at extremes of human performance. In connection with this, although soccer is considered to be a long-duration exercise [16, 17], it is well known that matches are won in high intensity, short burst efforts such as sprinting or jumping. Therefore, in addition to technical and tactical skills, muscle strength and “explosive” leg power are very important factors that contribute toward achieving successful results during elite soccer competitions. ACTN3 is vital in producing high-power and high-velocity muscle compression [16].
Due to the lack of data according to ACTN3 R577X polymorphism in Iranian athletes, in this study, we aimed to examine the associations between ACTN3 polymorphisms and athletic performance in a group of elite soccer players and compare their genotypic distributions with nonathletic controls.
2. Methods
Study participants
A total of 290 participants comprising of 90 elite male soccer players as case group and 200 unrelated healthy males as control group were recruited in this case–control study. All of the individuals in case group had represented Iran in their sport at the international level. The individuals in control group were volunteer males recruited from the Genetics Research Center (GRC). All the participants in this study were all of Iranian ancestry to overcome any potential issues with population stratification. Composed written assent was acquired from all participants under conventions affirmed by the institutional survey leading group of the GRC at the University of Social Welfare and Rehabilitation Sciences, Tehran, Iran.
Genotyping
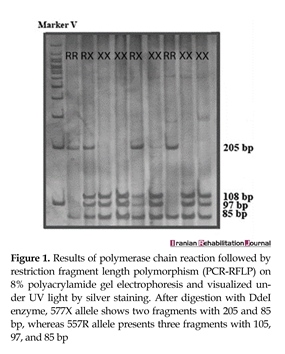
After obtaining written informed consent from all the participants, 10 mL of blood was collected in tubes containing ethylenediamine tetra-acetic acid (EDTA). Genomic DNA from all the samples was extracted according to the established protocols [18]. Exon 16 of ACTN3 was expanded through polymerase chain reaction (PCR) under standard conditions by using the following primers: forward 5′-CTGTTGCCTGTGGTAAGTGGG-3′, reverse 5′-TGGTCACAGTATGCAGGAGGG-3′. PCR products were digested with DdeI enzyme (New England Biolabs, Ipswich, MA, USA). The 577R and 577X alleles (CGA and TGA codons, respectively) were recognized by the presence (577X) or absence (577R) of a DdeI restriction site (Figure 1).
The α-actinin-3 structural protein is a fast skeletal muscle protein that is encoded by the α-actinin-3 gene (ACTN3, MIM#102574; chromosomal location: 11q13-q14) [1, 2]. ACTN3 has attracted considerable attention due to a common nonsense polymorphism (R577X) that influences muscular performance [3]. A loss-of-function nonsense mutation (C>T) at nucleotide position 1747 in the ACTN3 coding sequence is responsible for the transition of an arginine (R) to a premature stop codon (X) at residue 577 (R577X; dbSNPrs1815739).
This single nucleotide polymorphism is responsible for the distribution of two different types of ACTN3, both of which are common in general population [4]. The 577R allele is the normal, operational version of this gene, whereas 577X allele contains a changed sequence that prevents the production of a complete functional α-actinin-3 protein [5–7]. It is noteworthy that α-actinin-3 has relatively specialized expression and is limited to the fast, glycolytic muscle fibers that are responsible for quick force generation [6].
It is estimated that 16% of the individuals worldwide, or approximately one billion people, are deficient in α-actinin-3. Our previous study conducted on Iranian normal individuals showed that the allelic distribution of ACTN3 in Iranian population is closer to its distribution in the Caucasian population with 11% homozygosity for the common R577X polymorphism; this is concurrent with the route of migration of the ancient human population from the Iran Plateau toward Europe. ACTN3 knockout mouse model analysis has shown that α-actinin-3 deficiency brings about a “slowing” of fast fiber attributes conjoined with a dramatic fall in grip strength, muscle mass and fast 2B fiber size, a rise in muscle aerobic capacity, and slowing of muscle contractile properties [8].
In an Australian athlete cohort [9], highly significant associations between ACTN3 genotype and athletic performance was revealed. Both male and female elite sprint athletes had significantly higher frequencies of the 577R allele than that of control participants. Furthermore, in a number of studies conducted on European, Israeli, and North American athletes, it has been shown that 577XX genotype is underrepresented in sprint athletes and slightly overrepresented in endurance athletes compared with nonathletic controls [10–15]. These data, in addition to other recent studies, provide strong evidence that α-actinin-3 deficiency is detrimental to sprint and power performance.
The effect of ACTN3 genotype has primarily been studied in elite athletes as its effect on muscle can be most readily discernible at extremes of human performance. In connection with this, although soccer is considered to be a long-duration exercise [16, 17], it is well known that matches are won in high intensity, short burst efforts such as sprinting or jumping. Therefore, in addition to technical and tactical skills, muscle strength and “explosive” leg power are very important factors that contribute toward achieving successful results during elite soccer competitions. ACTN3 is vital in producing high-power and high-velocity muscle compression [16].
Due to the lack of data according to ACTN3 R577X polymorphism in Iranian athletes, in this study, we aimed to examine the associations between ACTN3 polymorphisms and athletic performance in a group of elite soccer players and compare their genotypic distributions with nonathletic controls.
2. Methods
Study participants
A total of 290 participants comprising of 90 elite male soccer players as case group and 200 unrelated healthy males as control group were recruited in this case–control study. All of the individuals in case group had represented Iran in their sport at the international level. The individuals in control group were volunteer males recruited from the Genetics Research Center (GRC). All the participants in this study were all of Iranian ancestry to overcome any potential issues with population stratification. Composed written assent was acquired from all participants under conventions affirmed by the institutional survey leading group of the GRC at the University of Social Welfare and Rehabilitation Sciences, Tehran, Iran.
Genotyping
After obtaining written informed consent from all the participants, 10 mL of blood was collected in tubes containing ethylenediamine tetra-acetic acid (EDTA). Genomic DNA from all the samples was extracted according to the established protocols [18]. Exon 16 of ACTN3 was expanded through polymerase chain reaction (PCR) under standard conditions by using the following primers: forward 5′-CTGTTGCCTGTGGTAAGTGGG-3′, reverse 5′-TGGTCACAGTATGCAGGAGGG-3′. PCR products were digested with DdeI enzyme (New England Biolabs, Ipswich, MA, USA). The 577R and 577X alleles (CGA and TGA codons, respectively) were recognized by the presence (577X) or absence (577R) of a DdeI restriction site (Figure 1).

Molecular genetic and statistical analysis
Chi-squared tests were used to compare genotypic distributions and allelic frequencies between the two groups and also to assess the presence of Hardy–Weinberg equilibrium. Data were analyzed using SPSS 18 (SPSS Inc, Chicago, IL, USA). P values less than 0.05 were considered to be statistically significant.
3. Results
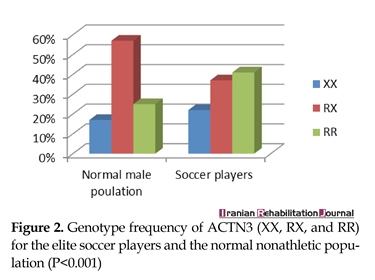
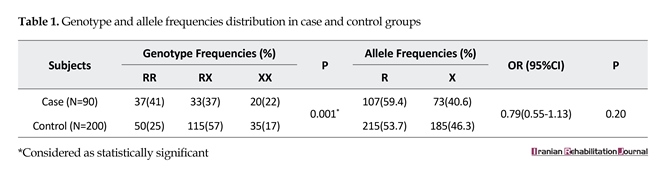
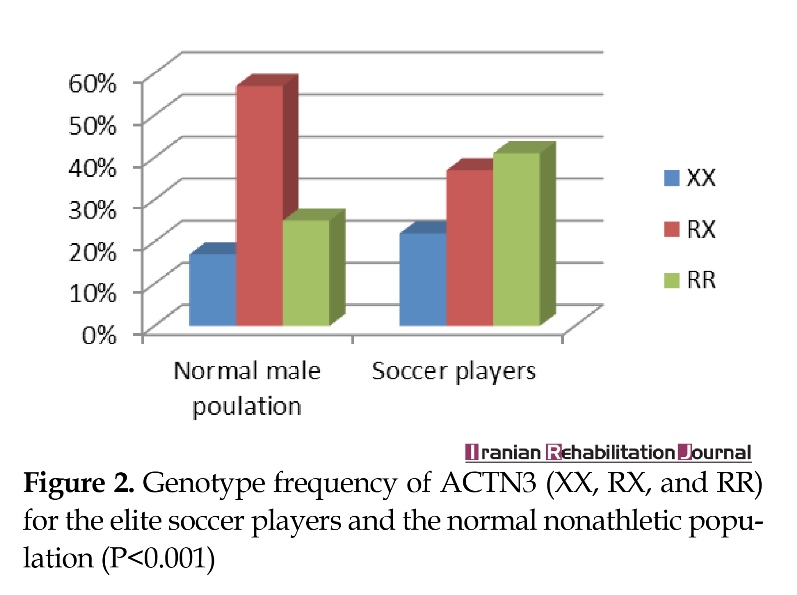
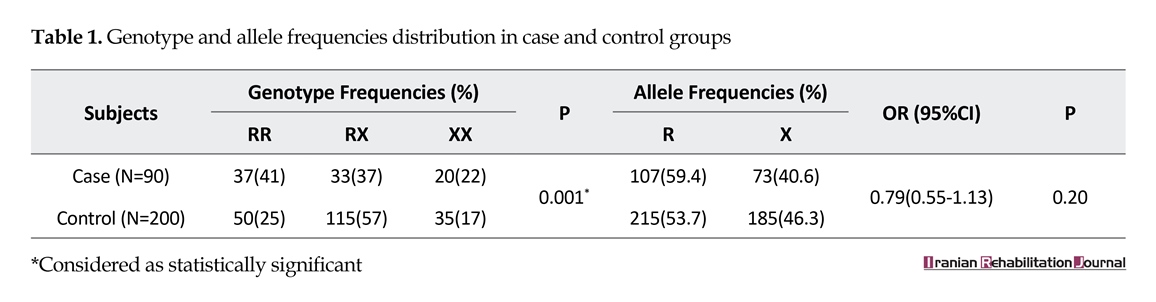
ACTN3 genotype data from the elite athlete and control groups are outlined in Table 1. There were significant diversities in genotypic frequency between the elite athlete group and the control group. Although the differences in allelic frequencies did not reach statistical significance, participants from elite athlete group did have a lower frequency of the 577RX genotype than that of 577RR and 577XX genotypes. The frequency of 577RX genotype was also lower in elite athlete group than that of control group (P<0.001). Genotypic frequencies of ACTN3 (XX, RX, and RR) for elite soccer players and normal nonathletic population (P<0.001) are shown in Figure 2.
Chi-squared tests were used to compare genotypic distributions and allelic frequencies between the two groups and also to assess the presence of Hardy–Weinberg equilibrium. Data were analyzed using SPSS 18 (SPSS Inc, Chicago, IL, USA). P values less than 0.05 were considered to be statistically significant.
3. Results
ACTN3 genotype data from the elite athlete and control groups are outlined in Table 1. There were significant diversities in genotypic frequency between the elite athlete group and the control group. Although the differences in allelic frequencies did not reach statistical significance, participants from elite athlete group did have a lower frequency of the 577RX genotype than that of 577RR and 577XX genotypes. The frequency of 577RX genotype was also lower in elite athlete group than that of control group (P<0.001). Genotypic frequencies of ACTN3 (XX, RX, and RR) for elite soccer players and normal nonathletic population (P<0.001) are shown in Figure 2.
4. Discussion
To the best of our knowledge, this is the first study to demonstrate a higher proportion of the ACTN3 577RR and 577RX genotypes in elite level speed-power athletes from Iran compared with the normal nonathletic population. Our findings revealed that Iranian elite athletes had a lower frequency of the heterozygous RX genotype (37% vs. 57%) compared with controls. Elite athletes had higher frequencies of RR and XX genotypes (41% and 25%, respectively) than that of control participants (22% and 17%, respectively) (P<0.001).
A strong positive association of ACTN3 577R polymorphism and athletes’ performance was first reported by Yang et al. in an Australian population [9] Since then, many studies have been performed in various populations and different athletic groups which have provided conclusive evidence of a correlation. Several cross-sectional studies to date have reliably shown that the 577XX genotype is underrepresented in sprint-related
To the best of our knowledge, this is the first study to demonstrate a higher proportion of the ACTN3 577RR and 577RX genotypes in elite level speed-power athletes from Iran compared with the normal nonathletic population. Our findings revealed that Iranian elite athletes had a lower frequency of the heterozygous RX genotype (37% vs. 57%) compared with controls. Elite athletes had higher frequencies of RR and XX genotypes (41% and 25%, respectively) than that of control participants (22% and 17%, respectively) (P<0.001).
A strong positive association of ACTN3 577R polymorphism and athletes’ performance was first reported by Yang et al. in an Australian population [9] Since then, many studies have been performed in various populations and different athletic groups which have provided conclusive evidence of a correlation. Several cross-sectional studies to date have reliably shown that the 577XX genotype is underrepresented in sprint-related
athletes [9, 12, 13] or is associated with inferior performance [19]. Two recent meta analyses [20, 21] and a revision [22] of the literature with regards to the effect of ACTN3 on athletic performance indicate that the 577RR genotype is affiliated with sprint-power athletes among Caucasians. A large cohort study of about 300 elite Japanese athletes indicated that the ACTN3 577RX genotype was associated with sprint-power performance among track and field contestants [23].
A study on Turkish male soccer players revealed that the frequencies of ACTN3 genotypes were 20, 36, and 44 for XX, RX, and RR, respectively, which agrees with the results of this study for each genotype [24]. In the Turkish study, an allelic frequency was found to be 31 (62%) and 19 (38%) for R and X alleles of ACTN3, respectively, which is similar to our findings (107 (59.4%) R and 73 (40.6%) X alleles). Similar results were obtained in Russian football players in which 240 male athletes were examined among which 46.25% showed the RR genotype [15].
Santiago et al. analyzed the periodicity of the ACTN3 genotype in 60 top-level professional soccer players from Brazil and showed that the distribution of 577RR and 577RX genotypes (48.3% and 36.7%, respectively) was notably higher and lower, as in the order, than those in controls (n=123; 28.5% and 53.7%, respectively) (p=0.041) [10]. The result of this study is also in accordance with the Brazilian study [10]. Another recent study by Pimenta et al., in which 37 professional soccer players were examined, showed a prevalence of 40.5% for the RR genotype [25], which agrees with our results.
The results of this study indicate that the ACTN3 RR genotype is overrepresented in Iranian elite male soccer players compared to control participants. Moreover, we showed that the distribution of the ACTN3 genotypes and alleles in the Iranian population is similar to those observed in several reported groups of Russian, Brazilian, and Turkish populations [10, 24, 26]. There were no statistically significant differences noted in the allelic frequencies between elite soccer players and control group participants, though sample size limitations prevented a conclusive analysis in this regard.
The ACTN3 discoveries reported here broaden past reports demonstrating higher 577RR genotype recurrence in elite speed/power-oriented athletes and competitors than that of normal participants. This supports to the speculation that α-actinin-3 protein deficiency (XX genotype) impairs muscle performance. The ACTN3 RR genotype, alone or in combination with additional polymorphisms such as the angiotensin converting enzyme insertion/deletion genotype, may be useful predictors of athletic performance at the elite level.
Acknowledgments
We are deeply grateful to the participants of this study, and we greatly appreciate Dr. Mohammad Dadkan, Dr. Amin Norouzi, Dr. Abdol Reza Allahverdi, and Mr. Alireza Mansourian for their collaboration which made this study possible. This project has not received any financial assistance. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Conflict of Interest
The authors declared no conflicts of interest.
References
[1]Beggs AH, Byers TJ, Knoll JH, Boyce FM, Bruns GA, Kunkel LM. Cloning and characterization of two human skeletal muscle alpha-actinin genes located on chromosomes 1 and 11. Journal of Biological Chemistry. 1992; 267(13):9281-8. PMID: 1339456
[2]MacArthur DG, North KN. A gene for speed? The evolution and function of actinin-3. BioEssays. 2004; 26(7):786–95. doi: 10.1002/bies.20061
[3]Rakus D, Mamczur P, Gizak A, Dus D, Dzugaj A. Colocalization of muscle FBPase and muscle aldolase on both sides of the Z-line. Biochemical and Biophysical Research Communications. 2003; 311(2):294–9. doi: 10.1016/j.bbrc.2003.09.209
[4]Fattahi Z, Najmabadi H. Prevalence of ACTN3 (the athlete gene) R577X polymorphism in Iranian population. Iranian Red Crescent Medical Journal. 2012; 14(10):617-22. PMCID: PMC3518978
[5]Vainzof M, Costa C, Marie S, Moreira E, Reed U, Passos-Bueno M, et al. Deficiency of α-actinin-3 (ACTN3) occurs in different forms of muscular dystrophy. Neuropediatrics. 1997; 28(04):223–8. doi: 10.1055/s-2007-973704
[6]Mills M. Differential expression of the actin-binding proteins, alpha-actinin-2 and -3, in different species: implications for the evolution of functional redundancy. Human Molecular Genetics. 2001; 10(13):1335–46. doi: 10.1093/hmg/10.13.1335
[7]Ahmetov II, Druzhevskaya AM, Astratenkova IV, Popov DV, Vinogradova OL, Rogozkin VA. The ACTN3 R577X polymorphism in Russian endurance athletes. British Journal of Sports Medicine. 2010; 44(9):649-52. doi: 10.1136/bjsm.2008.051540
[8]MacArthur DG, Seto JT, Chan S, Quinlan KG, Raftery JM, Turner N, et al. An actn3 knockout mouse provides mechanistic insights into the association between α-actinin-3 deficiency and human athletic performance. Human Molecular genetics. 2008; 17(8):1076-86. doi: 10.1093/hmg/ddm380
[9]Yang N, MacArthur DG, Gulbin JP, Hahn AG, Beggs AH, Easteal S, et al. ACTN3 genotype is associated with human elite athletic performance. The American Journal of Human Genetics. 2003; 73(3):627–31. doi: 10.1086/377590
[10]Santiago C, Gonzalez-Freire M, Serratosa L, Morate FJ, Meyer T, Gomez-Gallego F, et al. ACTN3 genotype in professional soccer players. British Journal of Sports Medicine. 2007; 42(1):71–3. doi: 10.1136/bjsm.2007.039172
[11]Druzhevskaya AM, Ahmetov II, Astratenkova IV, Rogozkin VA. Association of the ACTN3 R577X polymorphism with power athlete status in Russians. European Journal of Applied Physiology . 2008; 103(6):631–4. doi: 10.1007/s00421-008-0763-1
[12]Niemi A-K, Majamaa K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. European Journal of Human Genetics . 2005; 13(8):965–9. doi: 10.1038/sj.ejhg.5201438
[13]Papadimitriou I, Papadopoulos C, Kouvatsi A, Triantaphyllidis C. The ACTN3 gene in elite greek track and field athletes. International Journal of Sports Medicine. 2008; 29(4):352–5. doi: 10.1055/s-2007-965339
[14]Roth SM, Walsh S, Liu D, Metter EJ, Ferrucci L, Slavotinek AM. The ACTN3 R577X nonsense allele is under-represented in elite-level strength athletes. European Journal of Human Genetics . 2007; 16(3):391–4. doi: 10.1038/sj.ejhg.5201964
[15]Eynon N, Duarte JA, Oliveira J, Sagiv M, Yamin C, Meckel Y, et al. ACTN3R577X Polymorphism and Israeli top-level athletes. International Journal of Sports Medicine. 2009; 30(9):695–8. doi: 10.1055/s-0029-1220731
[16]Pimenta EM, Coelho DB, Veneroso CE, Barros Coelho EJ, Cruz IR, Morandi RF, et al. Effect of ACTN3 gene on strength and endurance in soccer players. Journal of Strength and Conditioning Research. 2013; 27(12):3286–92. doi: 10.1519/jsc.0b013e3182915e66
[17]Bangsbo J. The physiology of soccer--with special reference to intense intermittent exercise. Acta Physiologica Scandinavica. Supplementum. 1994; 619:1-55. PMID: 8059610
[18]Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Research. 1988; 16(3):1215–1215. doi: 10.1093/nar/16.3.1215
[19]Moran CN, Yang N, Bailey MES, Tsiokanos A, Jamurtas A, MacArthur DG, et al. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. European Journal of Human Genetics . 2006; 15(1):88–93. doi: 10.1038/sj.ejhg.5201724
[20]Alfred T, Ben-Shlomo Y, Cooper R, Hardy R, Cooper C, Deary IJ, et al. ACTN3 genotype, athletic status, and life course physical capability: Meta-analysis of the published literature and findings from nine studies. Human Mutation. 2011; 32(9):1008–18. doi: 10.1002/humu.21526
[21]Ma F, Yang Y, Li X, Zhou F, Gao C, Li M, et al. The association of sport performance with ACE and ACTN3 genetic polymorphisms: A systematic review and meta-analysis. PLoS ONE. 2013; 8(1):54685. doi: 10.1371/journal.pone.0054685
[22]Eynon N, Hanson ED, Lucia A, Houweling PJ, Garton F, North KN, et al. Genes for elite power and sprint performance: ACTN3 leads the way. Sports Medicine . 2013; 43(9):803–17. doi: 10.1007/s40279-013-0059-4
[23]Mikami E, Fuku N, Murakami H, Tsuchie H, Takahashi H, Ohiwa N, et al. ACTN3 R577X genotype is associated with sprinting in elite Japanese athletes. International Journal of Sports Medicine. 2013; 35(02):172–7. doi: 10.1055/s-0033-1347171
[24]Ulucan K, Sercan C, Biyikli T. Distribution of angiotensin-1 converting enzyme insertion/deletion and & alpha-actinin-3 codon 577 polymorphisms in Turkish male soccer players. Genetics & Epigenetics. 2015; 7:1-4. doi: 10.4137/geg.s31479
[25]Pimenta EM, Coelho DB, Cruz IR, Morandi RF, Veneroso CE, de Azambuja Pussieldi G, et al. The ACTN3 genotype in soccer players in response to acute eccentric training. European Journal of Applied Physiology . 2011; 112(4):1495–503. doi: 10.1007/s00421-011-2109-7
[26]Egorova ES, Borisova AV, Mustafina LJ, Arkhipova AA, Gabbasov RT, Druzhevskaya AM, et al. The polygenic profile of Russian football players. Journal of Sports Sciences. 2014; 32(13):1286–93. doi: 10.1080/02640414.2014.898853
A study on Turkish male soccer players revealed that the frequencies of ACTN3 genotypes were 20, 36, and 44 for XX, RX, and RR, respectively, which agrees with the results of this study for each genotype [24]. In the Turkish study, an allelic frequency was found to be 31 (62%) and 19 (38%) for R and X alleles of ACTN3, respectively, which is similar to our findings (107 (59.4%) R and 73 (40.6%) X alleles). Similar results were obtained in Russian football players in which 240 male athletes were examined among which 46.25% showed the RR genotype [15].
Santiago et al. analyzed the periodicity of the ACTN3 genotype in 60 top-level professional soccer players from Brazil and showed that the distribution of 577RR and 577RX genotypes (48.3% and 36.7%, respectively) was notably higher and lower, as in the order, than those in controls (n=123; 28.5% and 53.7%, respectively) (p=0.041) [10]. The result of this study is also in accordance with the Brazilian study [10]. Another recent study by Pimenta et al., in which 37 professional soccer players were examined, showed a prevalence of 40.5% for the RR genotype [25], which agrees with our results.
The results of this study indicate that the ACTN3 RR genotype is overrepresented in Iranian elite male soccer players compared to control participants. Moreover, we showed that the distribution of the ACTN3 genotypes and alleles in the Iranian population is similar to those observed in several reported groups of Russian, Brazilian, and Turkish populations [10, 24, 26]. There were no statistically significant differences noted in the allelic frequencies between elite soccer players and control group participants, though sample size limitations prevented a conclusive analysis in this regard.
The ACTN3 discoveries reported here broaden past reports demonstrating higher 577RR genotype recurrence in elite speed/power-oriented athletes and competitors than that of normal participants. This supports to the speculation that α-actinin-3 protein deficiency (XX genotype) impairs muscle performance. The ACTN3 RR genotype, alone or in combination with additional polymorphisms such as the angiotensin converting enzyme insertion/deletion genotype, may be useful predictors of athletic performance at the elite level.
Acknowledgments
We are deeply grateful to the participants of this study, and we greatly appreciate Dr. Mohammad Dadkan, Dr. Amin Norouzi, Dr. Abdol Reza Allahverdi, and Mr. Alireza Mansourian for their collaboration which made this study possible. This project has not received any financial assistance. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Conflict of Interest
The authors declared no conflicts of interest.
References
[1]Beggs AH, Byers TJ, Knoll JH, Boyce FM, Bruns GA, Kunkel LM. Cloning and characterization of two human skeletal muscle alpha-actinin genes located on chromosomes 1 and 11. Journal of Biological Chemistry. 1992; 267(13):9281-8. PMID: 1339456
[2]MacArthur DG, North KN. A gene for speed? The evolution and function of actinin-3. BioEssays. 2004; 26(7):786–95. doi: 10.1002/bies.20061
[3]Rakus D, Mamczur P, Gizak A, Dus D, Dzugaj A. Colocalization of muscle FBPase and muscle aldolase on both sides of the Z-line. Biochemical and Biophysical Research Communications. 2003; 311(2):294–9. doi: 10.1016/j.bbrc.2003.09.209
[4]Fattahi Z, Najmabadi H. Prevalence of ACTN3 (the athlete gene) R577X polymorphism in Iranian population. Iranian Red Crescent Medical Journal. 2012; 14(10):617-22. PMCID: PMC3518978
[5]Vainzof M, Costa C, Marie S, Moreira E, Reed U, Passos-Bueno M, et al. Deficiency of α-actinin-3 (ACTN3) occurs in different forms of muscular dystrophy. Neuropediatrics. 1997; 28(04):223–8. doi: 10.1055/s-2007-973704
[6]Mills M. Differential expression of the actin-binding proteins, alpha-actinin-2 and -3, in different species: implications for the evolution of functional redundancy. Human Molecular Genetics. 2001; 10(13):1335–46. doi: 10.1093/hmg/10.13.1335
[7]Ahmetov II, Druzhevskaya AM, Astratenkova IV, Popov DV, Vinogradova OL, Rogozkin VA. The ACTN3 R577X polymorphism in Russian endurance athletes. British Journal of Sports Medicine. 2010; 44(9):649-52. doi: 10.1136/bjsm.2008.051540
[8]MacArthur DG, Seto JT, Chan S, Quinlan KG, Raftery JM, Turner N, et al. An actn3 knockout mouse provides mechanistic insights into the association between α-actinin-3 deficiency and human athletic performance. Human Molecular genetics. 2008; 17(8):1076-86. doi: 10.1093/hmg/ddm380
[9]Yang N, MacArthur DG, Gulbin JP, Hahn AG, Beggs AH, Easteal S, et al. ACTN3 genotype is associated with human elite athletic performance. The American Journal of Human Genetics. 2003; 73(3):627–31. doi: 10.1086/377590
[10]Santiago C, Gonzalez-Freire M, Serratosa L, Morate FJ, Meyer T, Gomez-Gallego F, et al. ACTN3 genotype in professional soccer players. British Journal of Sports Medicine. 2007; 42(1):71–3. doi: 10.1136/bjsm.2007.039172
[11]Druzhevskaya AM, Ahmetov II, Astratenkova IV, Rogozkin VA. Association of the ACTN3 R577X polymorphism with power athlete status in Russians. European Journal of Applied Physiology . 2008; 103(6):631–4. doi: 10.1007/s00421-008-0763-1
[12]Niemi A-K, Majamaa K. Mitochondrial DNA and ACTN3 genotypes in Finnish elite endurance and sprint athletes. European Journal of Human Genetics . 2005; 13(8):965–9. doi: 10.1038/sj.ejhg.5201438
[13]Papadimitriou I, Papadopoulos C, Kouvatsi A, Triantaphyllidis C. The ACTN3 gene in elite greek track and field athletes. International Journal of Sports Medicine. 2008; 29(4):352–5. doi: 10.1055/s-2007-965339
[14]Roth SM, Walsh S, Liu D, Metter EJ, Ferrucci L, Slavotinek AM. The ACTN3 R577X nonsense allele is under-represented in elite-level strength athletes. European Journal of Human Genetics . 2007; 16(3):391–4. doi: 10.1038/sj.ejhg.5201964
[15]Eynon N, Duarte JA, Oliveira J, Sagiv M, Yamin C, Meckel Y, et al. ACTN3R577X Polymorphism and Israeli top-level athletes. International Journal of Sports Medicine. 2009; 30(9):695–8. doi: 10.1055/s-0029-1220731
[16]Pimenta EM, Coelho DB, Veneroso CE, Barros Coelho EJ, Cruz IR, Morandi RF, et al. Effect of ACTN3 gene on strength and endurance in soccer players. Journal of Strength and Conditioning Research. 2013; 27(12):3286–92. doi: 10.1519/jsc.0b013e3182915e66
[17]Bangsbo J. The physiology of soccer--with special reference to intense intermittent exercise. Acta Physiologica Scandinavica. Supplementum. 1994; 619:1-55. PMID: 8059610
[18]Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Research. 1988; 16(3):1215–1215. doi: 10.1093/nar/16.3.1215
[19]Moran CN, Yang N, Bailey MES, Tsiokanos A, Jamurtas A, MacArthur DG, et al. Association analysis of the ACTN3 R577X polymorphism and complex quantitative body composition and performance phenotypes in adolescent Greeks. European Journal of Human Genetics . 2006; 15(1):88–93. doi: 10.1038/sj.ejhg.5201724
[20]Alfred T, Ben-Shlomo Y, Cooper R, Hardy R, Cooper C, Deary IJ, et al. ACTN3 genotype, athletic status, and life course physical capability: Meta-analysis of the published literature and findings from nine studies. Human Mutation. 2011; 32(9):1008–18. doi: 10.1002/humu.21526
[21]Ma F, Yang Y, Li X, Zhou F, Gao C, Li M, et al. The association of sport performance with ACE and ACTN3 genetic polymorphisms: A systematic review and meta-analysis. PLoS ONE. 2013; 8(1):54685. doi: 10.1371/journal.pone.0054685
[22]Eynon N, Hanson ED, Lucia A, Houweling PJ, Garton F, North KN, et al. Genes for elite power and sprint performance: ACTN3 leads the way. Sports Medicine . 2013; 43(9):803–17. doi: 10.1007/s40279-013-0059-4
[23]Mikami E, Fuku N, Murakami H, Tsuchie H, Takahashi H, Ohiwa N, et al. ACTN3 R577X genotype is associated with sprinting in elite Japanese athletes. International Journal of Sports Medicine. 2013; 35(02):172–7. doi: 10.1055/s-0033-1347171
[24]Ulucan K, Sercan C, Biyikli T. Distribution of angiotensin-1 converting enzyme insertion/deletion and & alpha-actinin-3 codon 577 polymorphisms in Turkish male soccer players. Genetics & Epigenetics. 2015; 7:1-4. doi: 10.4137/geg.s31479
[25]Pimenta EM, Coelho DB, Cruz IR, Morandi RF, Veneroso CE, de Azambuja Pussieldi G, et al. The ACTN3 genotype in soccer players in response to acute eccentric training. European Journal of Applied Physiology . 2011; 112(4):1495–503. doi: 10.1007/s00421-011-2109-7
[26]Egorova ES, Borisova AV, Mustafina LJ, Arkhipova AA, Gabbasov RT, Druzhevskaya AM, et al. The polygenic profile of Russian football players. Journal of Sports Sciences. 2014; 32(13):1286–93. doi: 10.1080/02640414.2014.898853
Article type: Original Research Articles |
Subject:
Genetics
Received: 2016/10/11 | Accepted: 2017/01/28 | Published: 2017/08/1
Received: 2016/10/11 | Accepted: 2017/01/28 | Published: 2017/08/1
Send email to the article author

Copyright © The Author(s);
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC-By-NC 4.0), which permits use, distribution, and reproduction in any medium, provided the original work is properly cited and is not used for commercial purposes
Contact Information
